![]()

![]()



透過基因研究發現,靈芝免疫調節蛋白質(FIP-gts)是藉由胺端之胺基酸,形成雙體構形結合到淋巴球 T 細胞上,造成細胞凝集,使得 T 細胞活化而形成一連串之訊息傳遞,並促進分泌各種細胞激素來達成免疫調節之功能。
文/許瑞祥
◎本篇原載於1998年10月《健康靈芝》第2期12~15頁
靈芝自古為我國吉祥如意的象徵。靈芝的子實體則為扶正固本的珍貴藥材。至1970年代靈芝經由現代化科技被大量人工栽培後,藉由西方藥理學的研究模式,重新探討靈芝的藥用價值時,褪去了傳說中靈芝的神秘面紗,也更印證了古籍中記載靈芝具有安神、補中、益心氣、補肝氣、益精氣、堅筋骨、利水道、利腎氣、久食輕身不老等作用。
經由過去二十年的實驗動物研究證實,靈芝的萃取物中具有鎮靜、鎮痛、鎮咳、平喘、強心、保肝、降血糖、降血脂、降血壓、抗過敏、抗發炎、抗腫瘤、抗衰老、抗疲勞、免疫增強等活性,而受到各國研究人員的肯定與重視,成為延遲衰老、預防慢性疾病的寄託。
靈芝屬是屬於真菌界(Myceteae),無鞭毛菌門(Amastigomycota),擔子菌綱(Basidiomycetes),無蕈褶目(Aphyllophorales),多孔菌科(Polyporaceae)。靈芝屬的主要特徵,為其具有雙層細胞壁的擔孢子。
目前靈芝屬中在文獻中雖然有300種左右,但在藥理和臨床的研究中只有靈芝(G. lucidum)、松杉靈芝(G. tsugae) 、薄樹靈芝(G. capense)、狹長孢靈芝(G. boninense)、無柄靈芝(G. resinaceum)等紅色的靈芝和紫芝(G.sinense)、日本靈芝(G. japonicum)等紫色的靈芝與樹舌靈芝(G. applanatum)等少數的幾個特定種類的靈芝被研究與利用【1】。
在過去靈芝的藥理研究中,抗癌活性是最早被發現的,而靈芝的多醣體被認為是抗癌活性的主要成分,但是多醣體並不能直接殺死或抑制癌細胞,而是藉由提高免疫能力來間接表現其活性,除了可以增強 T 細胞 DNA polymerase-α活性促進其 DNA 之合成而活化 T 細胞外,並可增強自然殺手細胞之能力【2】。
此外,對於單核球的巨噬細胞的作用,會使其表現出吞噬能力增強及促進一些抑制腫瘤生長的細胞激素如:間白細胞素(IL-2、IL-4)、干擾素(INF-γ)、腫瘤壞死因子(TNF-α)等的合成與釋放【3】。因此藉由強化自然殺手細胞和巨噬細胞直接攻擊不正常的腫瘤細胞,達到防癌、抗癌之效果。
靈芝多醣體具有抑制腫瘤活性,其主要作用是藉由提升自體免疫能力來達到抗病目的,因此藉由提供訊號而啟動原本已有的能力,相當符合傳統醫學所謂「扶正固本」、「扶正祛邪」的觀念。
此作用相當類似於西方學者所謂「生物反應修飾劑(biological response modifier, BRM)」的概念,經由增強或調控機體的免疫功能,減輕致病因素對人體的損傷,而提高人體抗病能力,達到治病、防病的目的【4】。
這些物質與一般天然抗癌藥物作用機制不同,一般天然抗癌藥物多是抑制細胞之DNA、 RNA及蛋白質合成,而BRM【5】通常會結合到細胞表面專一受體,經由一系列訊息傳遞,例如:間白細胞素(IL-2、IL-4)、干擾素(NF-γ)、腫瘤壞死因子(TNF-α)等細胞激素之增加【6-8】,或引發抗體與補體的產生,進而達到提昇免疫能力,這些BRM通稱為「免疫增強劑」。
免疫增強劑包括不同來源,第一類是從微生物及天然物分離得來,天然物中 BCG【9-12】已被開發應用,OK432【13】、Lentinan【14】則是從微生物中純化而得到;第二類是人工合成物如:muramyl dipeptide【15】;第三類是由抗癌藥物中發現的如:cyclophosphamide【16】、adriamycin【17】;第四類則為目前發現之真菌類免疫調節蛋白質。
靈芝具有免疫調節蛋白質,在1989年日本學者Kino等人發表自 G. lucidum 菌絲中分離而得的小分子蛋白質 LZ-8 獲得證實【18】,LZ-8 的分子量約為13k,具有免疫調節和促進細胞有絲分裂的作用。
LZ-8 的發現使靈芝研究進入另一個全新的領域,同時也使靈芝的免疫調節作用再次受到注意。然而有趣的是,在一些同屬於擔子菌綱的真菌中,從子實體中可純化出一些外源凝集素(lectin),有些會促進淋巴球的增殖;相反的,有些會抑制淋巴球的增殖,包括洋菇(Agaricus bisporus),分子量為64K,由四個次單元體構成,會抑制細胞增殖。
野生洋菇(Agaricus compestris)分子量為4K、草菇(Volvariella volvacea)分子量為13K、Aleuria anrantia分子量為37K、Psathyrella velutina 分子量為40K、Lactarius delicious 分子量為37K,這些外源凝集素皆具有凝集細胞之能力。
一般而言,在蕈類中外源凝激素上所結合的醣類多不相同,猴頭菇(Hericium erinaceum)上的醣類是屬於sialic acid結合;Laetiporus sulfurens 是屬於 N-acetyl-galactosamine 結合。這些醣類可能促進細胞表面受體的結合,但其餘生理活性尚未深入探討。
外源凝集素於醫學上報導具有免疫調節(immunomodulatory)及保護肝臟(hepatoprotective)活性,可能是由於共有刺激細胞增生、促進釋放出些細胞激素(cytokines)及清除自由基(free radical)的功能。
由mistletoe分離出的外源凝集素,其為一種 β-galactoside 專一性的外源凝集素,不論體外(in vitro)或體內(in vivo)中皆會促進細胞激素增生。在Balb/C-老鼠的試驗中,對於腫瘤移植(tumor colonization)有抑制作用,並可促進人類腫瘤壞死因子(TNF-α)、間白細胞素(IL-l)的增加,具有免疫調節能力。
經基因重組的外源凝集素共有β-galactoside結合活性,可明顯抑制 autoimmune encephalomyelitis。Bolesatine 其蛋白質濃度為0.01微克/毫升時,有最強的細胞增生活性。大部分這類外源凝集素具有凝集細胞的作用,但其功能尚未清楚。
外源凝集素對於免疫調節功能方面,不同外源凝集素與不同抗體或細胞結合能力不同,如:Agaricus bisporus 會與免疫球蛋白IgA結合;bovine conglutinin 與 IgG 結合;L-selectin 促使嗜中性白血球(neutrophil)凝集;C. albicans 所純化得 manoprotein 其可促進人類單核球細胞(monocytes)增生。
lymphotoxin(淋巴毒蛋白),是屬於一種 lymphokine(細胞激素),有延遲性過敏反應作用。Ruddle 及Waksman 提到進行老鼠試驗中,當抗原存在時,淋巴球 T 細胞會釋放一種物質,引發淋巴細胞活化,會將相同基因型(synergenic)的老鼠胚胎纖維細胞殺死。Granger 等人稱這些具有細胞毒素者為「淋巴毒蛋白」。
如 phytohemagglutinin(PHA)是一種T細胞刺激原(mitogen),淋巴毒蛋白質會增加淋巴球對 3H-thymidine 之吸收及淋巴球細胞之增生。此類淋巴毒蛋白質的生物活性常伴隨著細胞激素的產生,二者具協力性遺傳毒性,特別是干擾素(IFN-γ)的產生。
淋巴毒蛋白質對標的物之影響,尚未完全了解,而淋巴毒蛋白所扮演之角色仍具有爭議,可能被當作 helper T 細胞所釋放之物質。但又不易見到促進抗體增加,淋巴毒蛋白質可能在發炎反應中扮演重要角色,對腫瘤或被侵害的組織具有防禦能力。
1990年代初,作者等開始有關靈芝來源的免疫調節蛋白質與其基因序列之分析。1994年我們由松杉靈芝(G. tsugae)菌絲體中純化出一免疫調節蛋白質,命名為 FIP-gts (fungal immunomodulatory protein- gts),分子量約為13K,含有少量的醣類,其不但可以促進人類周邊淋巴球細胞(human peripheral lymphocytes),也可以促進小白鼠脾臟細胞增生。
由細胞攝入3H-thymidine 之量與蛋白質濃度作圖,可得一鐘形圖形,在濃度為 5 μg/ml 時,對人類周邊淋巴球細胞可達到最高之增殖作用,利用 RT-PCR 方法也可證實其可促進細胞激素如間白細胞素(IL-2、IL-4)、干擾素(INF-γ)、腫瘤壞死因子(TNF-α)之表現。
免疫調節蛋白質(FIP-gts)經由蛋白酵素水解後,利用自動胺基酸分析儀(automatic protein sequencer)分析其胺基酸序列,可知含有110個胺基酸(圖一),利用分子遺傳技術經由序列比對,發現FIP-gts之胺基酸序列與日本人由靈芝(G. lucidum)純化出之免疫調節蛋白質LZ-8有相同序列【19-20】。
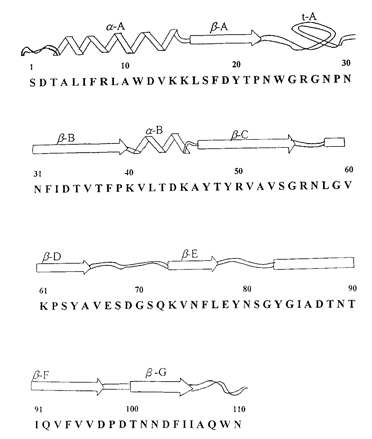
由目前發表之結果顯示,LZ-8 可以有效的抑制非肥胖糖尿病老鼠的自體免疫性第一型糖尿病之發生【21】,對 B 型肝炎表面抗原所引起的抗體亦有明顯的抑制作用,LZ-8 也可以結合T細胞及促進細胞激素之釋放【22-23】。
雖然我們從不同的靈芝菌種中,純化出與 LZ-8 具有相同之胺基酸序列,為了暸解在其他靈芝屬中是否也存在著此免疫調節基因,故我們也篩選多種不同菌種的靈芝,如:G. lucidum RSH RZ-TW、G. tropicum RSH 1111-TW、G. microsporum RSH 0821-TW 及 G. formosanum RSH 0109-TW。結果令人意外的是,在這些靈芝中的確存在此免疫調節基因,而且也具有相似的基因序列。
除此之外,也抽取數十種不同菌種靈芝的基因,利用 PCR 增幅方法來篩選是否存在有此免疫調節蛋白質基因,很意外地,在將近五十株不同菌種的靈芝中,均可以得到大小相似的基因產物,至於其在真菌中扮演何種角色則尚不明暸。
然而很有趣地,在一些非靈芝屬的真菌類中也可純化出免疫調節蛋白質,例如:在菇類中可由金針菇【24】及草菇純化出一分子量約為 13K 的免疫調節蛋白質,與從靈芝純化之 FIP-gts 大小差不多。
在其他菇類中除了金針菇及草菇外,則不存在此蛋白質,且經由胺基酸序列分析三者共有51個相同胺基酸,具有很高的相似性(圖二),更有趣的是此三種免疫調節蛋白質均含有與免疫球蛋白重鏈的可變區域有相當之相似性。
因此,我們可以推論由靈芝、金針菇及草菇所純化出之免疫調節蛋白質在演化上可能都是來自同一祖先,故我們也是國內最早發現不同的菌種中,也確實存在此免疫調節蛋白質基因,至於為什麼在靈芝屬中都有相同的序列及其在真菌類細胞中扮演何種角色?則必須再更深入的探討。

為了更進一步探討免疫調節蛋白質的作用機制,我們除了從松杉靈芝菌絲體中純化出此免疫調節蛋白質外,甚至將此免疫調節蛋白質利用遺傳工程技術,藉由大腸桿菌細胞大量地表現出來,並配合分子生物之刪除突變及定點突變技術,可以得到許多不同突變的突變蛋白質,用以探討其他生理活性與此蛋白質之活性區位及作用機制,這也是利用分生研究免疫調節蛋白質(FIP-gts)作用機制的首創成果。
經由免疫細胞增生活性分析可知,重組蛋白質(FIP-gts)與由菌絲體所純化之native FIP-gts有相同活性,在濃度為5μg/ml時有最高增殖活性。以膠體過濾方法可證實,免疫調節蛋白質(FIP-gts)在生理狀態下是以雙體形式存在。
同時由實驗結果顯示,若將胺基端刪除13胺基酸,突變蛋白質除了喪失凝集細胞之能力外,同時也無法促進細胞激素間白細胞素(IL-2、IL-4)、干擾素(INF-γ)、腫瘤壞死因子(TNF-α)等之分泌與免疫增生活性有關。
綜合上述結果證實,免疫調節蛋白質(FIP-gts)是藉由胺端之胺基酸形成雙體構形結合到淋巴球T細胞上,造成細胞凝集,使得T細胞活化而形成一連串之訊息傳遞,並促進分泌各種細胞激素來達成免疫調節之功能。
靈芝的免疫調節蛋白質除了有促進細胞增生外,也被發現對人類單核球細胞之免疫反應有抑制作用,同時也可以壓制人類淋巴球細胞對同種組織移植之反應,及延長同種異體之皮膚移植的生存時間。
在我們暸解免疫調節蛋白質之生理活性與作用機制後,希望將來能夠利用分子模型來合成此類小分子之有機化合物,應用於臨床研究上,並開發治療自體免疫疾病及用於器官移植之新的藥物。
我們從靈芝的免疫調節蛋白質 LZ-8 及基因的研發結果,開拓了另一個全新的領域。在免疫疾病束手無策的本世紀末,靈芝免疫調節蛋白質的發現,除了再次彰顯靈芝豐富的內涵外,以現代最新的實驗技術來檢視古老的靈芝扶正固本的作用時,怎不令人對大自然造物之神奇而讚嘆不已!
1. 許瑞祥,1993,靈芝概論,萬年出版社,140頁。
2. Lei LS, Lin ZB. 1991. Effect of Ganoderma polysaccharides on the activity of DNA polymerase α in spleen cells stimulated by alloantigens in mice in vitro. J. Beijing Medical University, 23(4): 329-333.
3. Lieu CW, Lee SS, Wang SY. 1992. The effect of Ganoderma lucidium on induction of differentiation in leukemic U937 cells. Anticancer Research, 12 (4): 1211-1215.
4. Mizuno T, Sakai T, Chihara G. 1995. Health foods and medicinal usages of mushrooms. Food Reviews International, 11(1) : 69-81.
5. Mihich E. 1986. Future perspectives for biological response modifiers: a viewpoint. Semin. Oncol. 13: 234-254.
6. Kampschmidt RF, Pulliam LA. 1975. Stimulation of antimicrobial activity in the rat with leukocyte endogenous mediator. J. Reticuloendothel Soc. 17: 162-169.
7. Dinarello CA. 1984. Interleukin-1. Rev. Infect Dis. 6: 51-95.
8. Dinarello CA, Cannon JG, Mier JW, Bernhein HA, LoPreste G. 1986. Multiple biological activities of human recombinant interleukin-l. J. Clin. Invest. 77: 1734-1739.
9. Baldwin RW, Byers VS. 1980. Immunoregulation by bacterial organisms and their role in the immunotherapy of cancer. In: Chedid L, Miescher PA, Mueller-Eberhard HJ (eds.): Immunostimulation. Berlin, Springer-Verlag, pp. 73-94.
10. Van’t Wout JW, Poell R, Van Furth R. 1992. The role of BCG/PPD-activated macrophages in resistance against systemic candidiasis in mice. Scand. J. Immunol. 36: 713-719.
11. Dubos RJ, Schaedler RW. 1957. Effects of cellular constituents of mycobacteria on the resistance of mice to heterologous infections. J. Exp. Med. 106: 719-726.
12. Sher NA, Chaparas SD, Greenberg LE, Bernard S. 1975. Effects of BCG, Corynebacterium parvam, and methanol-extraction residue in the reduction of mortality from Staphylococcus aureus and Candida albicans infections in immunosuppressed mice. Infect. Immun. 12: 1325-1330.
13. Shiraishi A, Mikami Y, Arai T. 1979. Protective effect of OK-432 ( a Streptococcal preparation) on experimental Candidiasis. Microbiol. Immunol. 23: 549-554.
14. Maeda YY, Chihara G. 1971. Lentinan, a new immuno-accelerator of cell mediated responses. Nature 229: 634-637.
15. Zidek Z, Frankova D, Masek K. 1993. Some cellular and pathophysiological correlates of the inflammatory effects of a synthetic immunomodulatory agent, muramyl dipeptide (MDP). Agents Actions. 38: 106-115.
16. Hengst JCD, Kempf RA. 1984. Immunomodulation by cyclophosphamide. In: Mitchell MS, Fahey JL. (eds.) : Clinics in Immunology and Allergy. Immune Suppression and Modulation, Vol. 4. London, W. B. Saunders, pp. 199-216.
17. Mihich E, Ehreke MJ, Ishizuka M. 1985. Immunomodulation by antibiotics. In Mihich E, Sakurai K. (eds.) : Biological responses in cancer, vol. 3, Immunomodulation by anticancer drugs. New York, Plenum Press, pp. 71-94.
18. Kino K, Yamashita A, Yamaoka K, Watanabe J, Tanaka S, Ko K, Shimizu K, Tsunoo H. 1989. Isolation and characterization of a new immunomodulatory protein, Ling Zhi-8 (LZ-8), from Ganoderma lucidum. J. Biol. Chem. 264: 472-478.
19. Murasugi A, Tanaka S, Komiyama N, Iwata N, Kino K, Tsunoo H, Sakuma S. 1991. Molecular cloning of a cDNA and a gene encoding an immunomodulatory protein, Ling Zhi-8, from a fungus, Ganoderma lucidum. J. Biol. Chem. 266: 2486-2493.
20. Tanaka S, Ko K, Kino K, Tsuchiya A, Yamashita A, Murasugi A, Sakuma S, Tsunoo H. 1989. Complete amino acid sequence of an immunomodulatory protein, Ling Zhi-8 (LZ-8). An immunomodulator from a fungus, Ganoderma lucidum, having similarity to immunoglobulin variable regions. J. Biol. Chem. 264: 16372-16377.
21. Kino K, Mizumoto K, Sone T, Yamaji T, Watanabe J, Yamashita A, Yamaoka K, Shimizu K, Ko K, Tsunoo H. 1990. An immunomodulating protein, Ling (Zhi-8) prevents insulitis in non-obese diabetic mice. Diabetologia 33: 713-718.
22. Miyasaka N, Inoue H, Totsuka T, Koike R, Kino K, Tsunoo H. 1992. An immunomodulatory protein, Ling Zhi-8, facilitates cellular interaction through modulation of adhesion molecules. Biochem. Biophys. Res. Commun. 186: 385-390.
23. Haak-Frendscho M, Kino K, Sone T, Jardieu P. 1993. Ling Zhi-8: A novel T cell mitogen induces cytokine production and upregulation of ICAM-1 expression. Cell Immunol. 150: 101-113.
24. Ko JL, Hsu CI, Lin RH, Kao CL, Lin JY. 1995. A new fungal immunomodulatory protein, FIP-fve isolated from the edible mushroom, Flammulina velutipes and its complete amino acid sequence. Eur. J. Biochem. 228: 244-249.
 Publication Date: Oct 3 2023Author: Ruey-Shyang Hseu, Michael Lee Publisher: Wu Ting Yao Studio, Taipei,...
Publication Date: Oct 3 2023Author: Ruey-Shyang Hseu, Michael Lee Publisher: Wu Ting Yao Studio, Taipei,...
COPYRIGHTS © Ganodermanews SINCE By 2015 ALL RIGHT RESERVED.
靈芝新聞網 版權所有 | 本網站內容作者享有其著作權,禁止侵害,違者必究
Powered by
ganodermanews.com
 最新期/2024年春季號
最新期/2024年春季號