![]()

![]()



專研靈芝三十多年的臺大教授許瑞祥,把靈芝產品和產業的演變,根據「吃出有效」的可靠性,分為1.0、2.0、3.0、4.0四個等級和階段,其所代表的是,靈芝自古至今被人們應用,在種源、生產、成分、功效等各層面,從「模糊不定」進步到「清楚穩定」的里程碑。產業和產品是相對應的。繼〔上篇〕介紹靈芝1.0~3.0產業和產品的特色之後,接下來的〔中篇〕將介紹以多糖與三萜為活性成分指標的靈芝4.0。你吃的靈芝是屬於4.0世代的嗎?
文.圖/許瑞祥(國立臺灣大學生化科技學系兼任教授)
成分決定功效。靈芝從第一個活性成分被發現後歷經三十年的探索,科學界從2000年開始,陸續發表有關靈芝高度純化成分的作用機制,用細胞或分子模式研究其作用的靶點與訊息傳遞途徑,精細描繪活性分子在癌化細胞內開啟或關閉各種導致細胞凋亡或自噬的結果,為靈芝抗癌作用提供堅實的理論基礎,讓靈芝能抗癌不再只是期待的願望,也為4.0版的靈芝產品和產業奠下基礎。
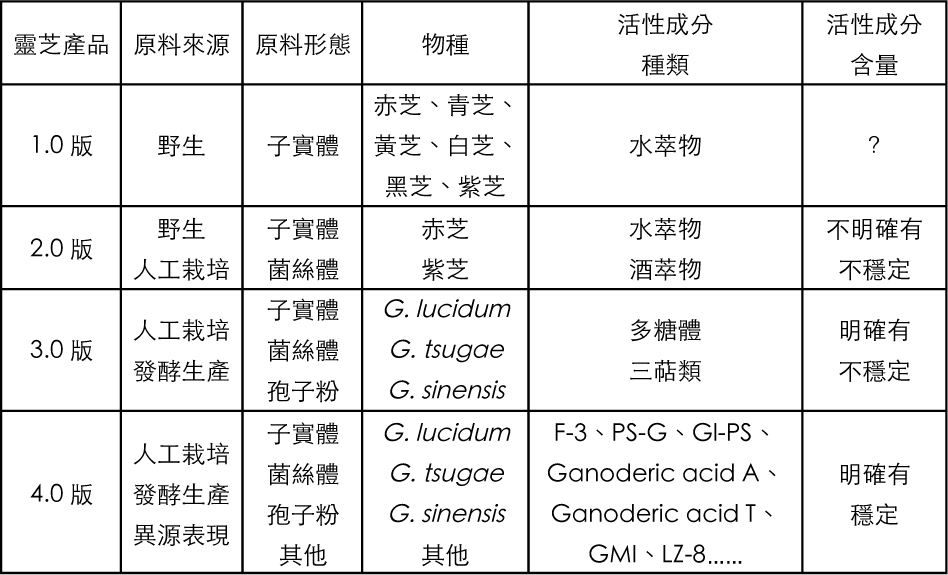
多糖體是大家最熟悉的靈芝活性成分。1971年Sasaki等從野生樹舌靈芝(G. applanatum)子實體中萃取具有抗肉瘤(S-180)細胞增生的活性成分,認為其活性來源是具有β(1→3)鍵結主鏈的葡萄糖聚合物,開啟了靈芝多糖體的研究。
1981年Usui等人發表從樹舌靈芝分離出兩種具有抗腫瘤活性的多糖體,皆為熱水可溶的β-D-glucan──具有β(1→3)鍵結主鏈與β(1→6)鍵結分支的葡聚糖──此β-D-glucan即為大家最熟悉的高分子多糖體活性構形的基本組成。
1984年水野卓發表靈芝(G. lucidum)子實體經熱水萃取、酒精沉澱、管柱層析後部分純化的活性多糖體,其中分子量1,050,000的β-D-glucan是「以β(1→3)鍵結為主鏈,含有分歧頻度4的β(1→6)鍵結,由六個糖連結的短支鍵構造」的抗腫瘤活性多糖體。
1985年Sone等人發表的報告,對靈芝子實體和菌絲體中經熱水或稀鹼溶液萃取而得的各種多糖體抗腫瘤活性進行比較。作者強調抗腫瘤活性強弱與其多糖體側鏈分支的頻度有關,被降低分支側鏈的多糖體,其抗腫瘤活性相對降低。本篇論文亦為比較子實體和菌絲體來源多糖體組成構形差異與抗腫瘤作用關係的代表作。
此時期所發表多糖的分離純化與組成分析技術日趨完整,對於抗腫瘤試驗的多糖體分子量大小、鍵結方式與組成構形差異已有較精確的描述。
1993年Wang等發表以松杉靈芝(G. tsugae)子實體來源的多糖體,經系列的分離與純化後進行抗腫瘤活性評估,在其所得的十四種水溶性多糖與十五種水不溶性多糖中,發現只有七種具有抗腫瘤(S-180)活性,皆由β(1→3)-D-glucan為主鏈,並且含有部分蛋白質的糖蛋白聚合物。
作者同時亦比較三種具有抗腫瘤活性的靈芝、樹舌靈芝、松杉靈芝間的多糖體組成與抗S-180肉瘤活性的評估。本研究使用三個物種為探討對象,彰顯不同物種間多糖組成的歧異,同時亦可瞭解在眾多多糖構形間,抗腫瘤活性會有極大的落差,更明確的界定何謂活性多糖。
在不同材料的研究報告間,所謂多糖體的含量、組成與效果,是無法互相推論或引用的,因此不同原料基礎所生產的靈芝產品功效自然也不能互相類比。
雖然已知靈芝活性多糖體是其抗腫瘤的重要有效成分,對於試驗動物的移植性腫瘤具有生長抑制作用,並可使腫瘤的重量減輕,延長寄主動物生存的時間,但在活體內使用的多糖體,是直接殺死癌細胞,還是經由激發寄主體內抗腫瘤機制間接殺死癌細胞,此時尚無定論。
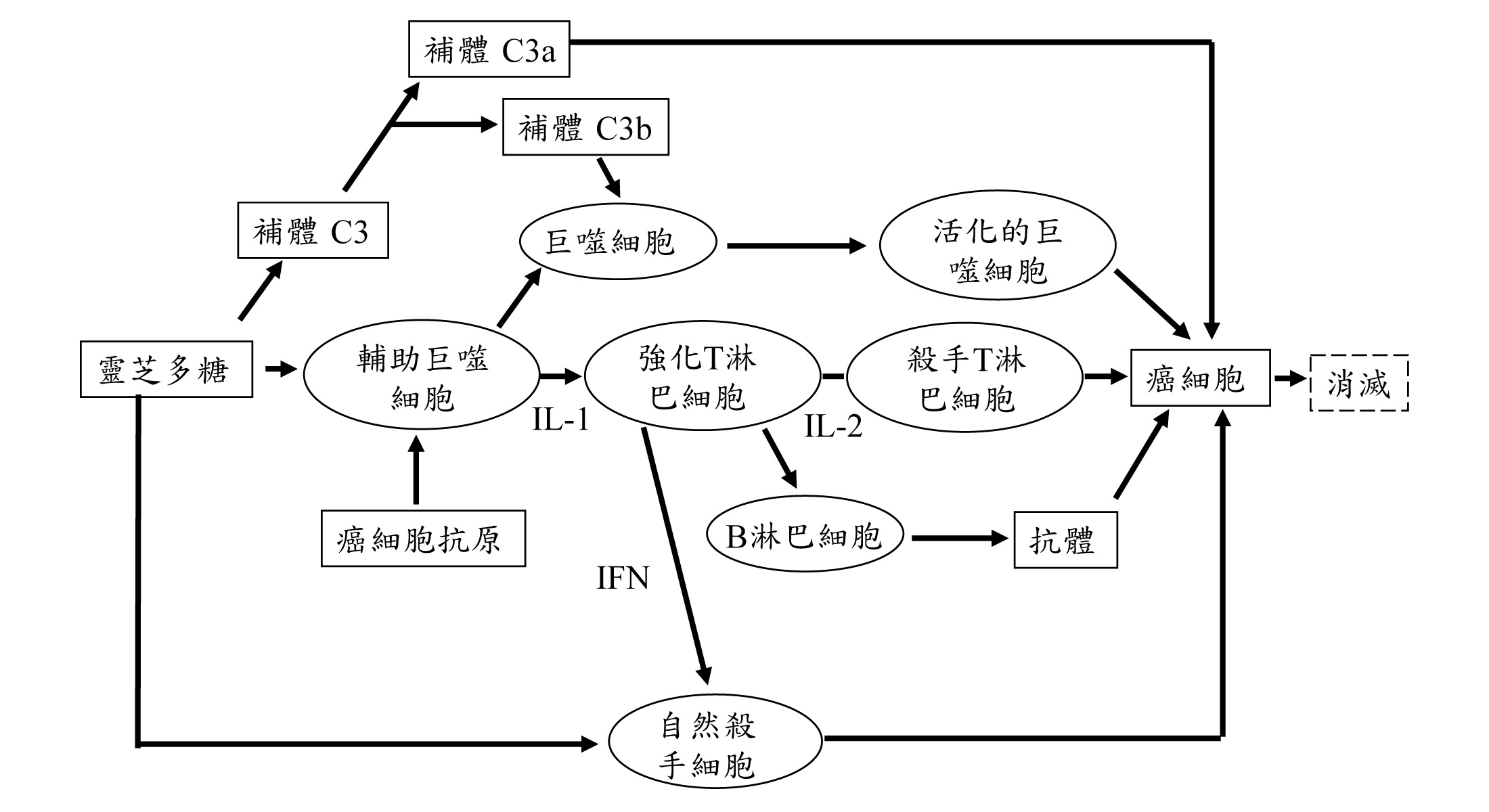
1997年王聲遠等發表靈芝多糖能抑制腫瘤細胞的增殖作用,是透過免疫系統中巨噬細胞和T淋巴細胞產生的TNF-α和IFN-γ協同作用的結果。
靈芝與免疫的研究成果有了連接之後,靈芝多糖體抗腫瘤的機制才被確認,靈芝與免疫系統研究的精彩序幕也隨之開啟,靈芝研究的模式亦從動物推展至細胞株,從腫瘤細胞推展至免疫細胞,從體內推展到體外。
2002年翁啟惠主導有關靈芝多糖體的系列研究,分析靈芝(G. lucidum)子實體經鹼萃取酸中和乙醇沉澱的水溶性粗萃取物,發現其碳水化合物組成以葡萄糖(58%)和甘露糖(15.5%)為主,並含有岩藻糖(9.7%)、半乳糖(9.3%)和木糖(5.4%)等醣類,同時該粗萃取物中亦含有約15.6%的蛋白質。
此粗萃取物經膠體濾管柱層析後,回收得到五個主要含醣的部分。以小鼠脾細胞增生試驗評估其主要活性表現的第三部分(F3),證明F3除了能促進脾細胞的增生外,亦可增加IL-1、IL-2和INF-γ的表現,並指出此高活性的F3組成是含有岩藻糖的醣蛋白。靈芝研究自此從基因體學進入蛋白質體學的領域,對於活性多糖體的組成與構形具備更精確的描述能力,成分精製的F3靈芝產品於是誕生。
在許先業等所發表的論文中證實,F3是經由與巨噬細胞表面TLR4受體的結合來啟動細胞內IL-1基因的表現。該研究對於IL-1基因於表現前的轉錄,F3調控後轉錄的pro-IL-1,以及IL-1的生成與分泌至胞外的過程,皆有清楚的瞭解,讓我們對於靈芝多糖體在細胞外如何經由特定TLR4受體來啟動胞內的基因表現,進而生成細胞激素後分泌到胞外參與免疫調節的過程,有更精確的認識。
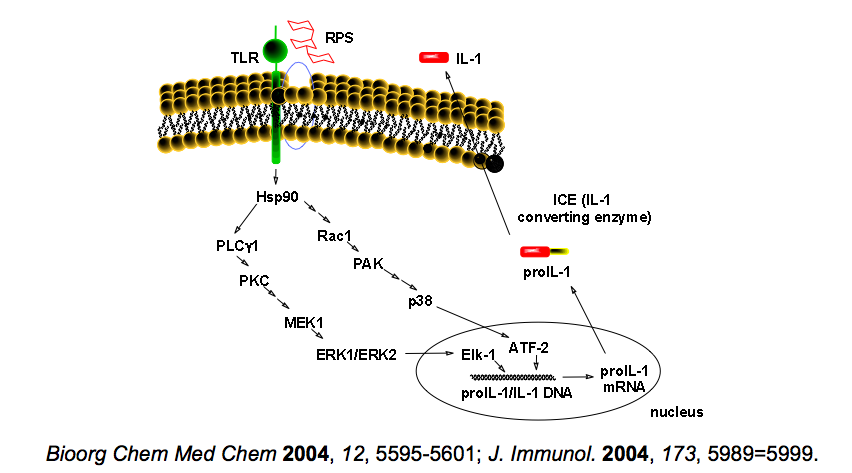
大約在同時期,另一組團隊亦發表由靈芝(G. lucidum)子實體經熱水萃取、酒精沉澱的粗多糖,經過膠體管柱層析與陰離子交換樹脂管柱分離純化所得的高分子多糖體,代號為PS-G,此 PS-G是由95%的多糖體和5%的胜肽組成的多糖肽。
2005年Lin等證實PS-G亦是經由樹突細胞表面的TLR-4受體將訊息傳入細胞內,能促進樹突細胞的成熟,進而合成並分泌細胞激素IL-12。IL-12能增強自然殺細胞和T細胞的噬菌能力,促使自然殺手細胞分泌INF-γ,還能誘導T細胞朝向第一型免疫反應分化,多糖體可經由細胞表面特定受體來啟動、調節免疫功能,再度獲得證實。
林志彬的研究團隊發表靈芝(G. lucidum)子實體經熱水提取、酒精沉澱、透析與去蛋白質等程序後所得多糖體,代號Gl-PS,此多糖經組成分析後,確認也是多糖肽,分子量為584,900,多糖和肽的比例為93.51:6.49。
其中多糖由鼠李糖(rhamnose)、木糖(xylose)、果糖(fructose)、半乳糖(galactose)、甘露糖(mannose)和葡萄糖以0.793:0.964:2.944:0.167:0.384:7.94比例組成,多肽的部分則由十七種胺基酸所組成。以Gl-PS處理樹突細胞後,不但能加速其成熟,同時可以誘導毒殺性T細胞生成。
2004年Shao證實Gl-PS能活化小鼠的B細胞和巨噬細胞,但不能活化T細胞。該研究以細胞表面「具有TLR-4受體」與「不具TLR-4受體」的B細胞和巨噬細胞互相交互驗證,再次確認靈芝多糖體是經由「與TLR-4受體結合後向胞內傳遞訊息」來達成調控免疫的目的。
靈芝多糖組份Gl-PS、PS-G和F3都是含有蛋白質的多糖肽,其研究來源不同,分離純化條件各異,但無論是B細胞、巨噬細胞或樹突細胞,皆是與TLR-4受體結合後開始啟動免疫訊息傳遞,由此結果可知,此類活性多糖肽的基本構造必定有其相似之處,而過去模糊的高分子多糖體,也逐步釐清到活性多糖肽的概念。
1982年Kubota等發現靈芝G. lucidum子實體的兩種新苦味成分,命名為ganoderic acid A和 B(靈芝酸A和B)。靈芝酸A分子式C30H44O7分子量516.3,是一種新型高度氧化的三萜類化合物。
1983年Toth等自靈芝G. lucidum菌絲體中分離靈芝酸T、S、R等六種新的三萜類化合物。
1990年Hirotani等發表以HPLC分析G. lucidum的菌絲體與子實體不同部位、不同生長階段的三萜類成分變化,發現有些靈芝酸(如靈芝酸A、B、H)僅存在於子實體,而靈芝酸T、S和R則是菌絲體與子實體都有。
1998年Min等從靈芝G. lucidum孢子中分離出的十二種萜類化合物,其中ganoderiol F和ganodermanontriol被發現具有抗HIV活性,靈芝酸β、lucidumol B和ganolucidic acid A也對HIV-1蛋白酶具有抑制活性,能控制愛滋病毒的複製,在當時被認為是世紀黑死病的救星。
靈芝在不同的生活史階段,不論是子實體、孢子或菌絲皆會產生特定的三萜類,呈現不同的生理功能。目前自靈芝G. lucidum和松杉靈芝G. taugae子實體、孢子和菌絲體所分離而得的三萜類化合物經鑑定者已達兩百種以上,其中以四環三萜化合物為主。
在這些構造相似且皆屬於高度氧化的羊毛固烷衍生物中,有的苦味很強,如酸性的三萜類化合物靈芝酸(ganoderic acid)和赤芝酸(lucidenic acid),有些則是淡而無味的。
這些靈芝特有的三萜類化合物經三十多年的藥理研究結果顯示具有:保肝作用、抗腫瘤、抗病毒增生、抑制組織胺釋放、抑制膽固醇合成、抑制血小板聚集、抗氧化作用、抗發炎作用、治療過敏性氣喘、預防骨質疏鬆症、改善下泌尿道症候群等功能,成為靈芝功能性產品開發的標靶。
開發含有特定三萜類組成與含量的產品,是「一定有效」的根本。以最早被發表的靈芝酸A為例,從早期的抑制組織胺釋放,到後來的抗癌細胞增殖與侵襲,抑制癌細胞訊息傳遞作用等,已成為靈芝產品功能性指標成分之一。
由於靈芝酸A只存在子實體萃取物裡,目前尚無法以人工合成方式取得,要想量產靈芝酸A就必須由大量子實體來萃取。選擇靈芝酸A高產量的靈芝品種,正確的栽培與管理方式,以及適當採收時期與庫存管理,都是提高產量降低成本的功課。
2016年Sakamoto等發表一種高特異性抗體(MAb12A)針對靈芝酸A進行專一性檢測,其偵測極限為6 ng/mL,能夠用在口服靈芝酸A後的血液裡追蹤其動力學的變化,也可以用於提昇靈芝產品質量控制的準確性和可靠性。
另一種在菌絲體也大量存在,能夠毒殺肝癌細胞的靈芝酸T,後續研究此靈芝酸T對於肺腺癌與子宮頸癌細胞的抑制作用時,發現它能使癌細胞的週期停滯在G1期,並可藉由降低粒線體膜電位、提高細胞內caspase-3和caspase-9的活性等機制,誘導癌細胞凋亡,成為標靶抗癌開發的選項。
靈芝酸T可以由靈芝菌絲體中萃取而得,因此開啟了菌種篩選、發酵生產的製程改良、回收分離技術的提昇的競賽。2012年Li等發表應用ADS-8樹脂的回收和再利用技術,提高靈芝菌絲體粗萃取物純化到最終靈芝酸T的回收率達72.2%,減少精製加工過程的損耗。
2012年Liu等將靈芝酸T 進行結構修飾後,產生更有效的小分子化合物,對子宮頸癌HeLa細胞的生長有更高的抑制效果,但是對正常細胞毒性更低,讓靈芝酸T成為開發癌細胞增殖抑製劑的重要原料。
2016年Zhang等發表以基因工程來調控同源的羊毛固醇合成酶(LS)基因複製數目,以及在靈芝菌絲體中的過度表達,結果顯示可以增加菌絲體中靈芝酸和羊毛固醇的含量,成分分析證實轉基因菌株靈芝酸A的含量提高3.2倍,羊毛固醇和麥角固醇的累積量也比對照菌株高2.3和1.4倍。
這項研究表明,同源合成酶基因的轉殖與過度表達的誘導可以促使羊毛固醇的大量積累,作為靈芝酸生物合成時的前趨物,增加靈芝酸產量,為菌絲體來源的靈芝酸生產成本的降低做出貢獻。
4.0版靈芝產品「一定有效」的特徵:
1. 具備至少一種可正確標示含量的靈芝活性成分。
2. 此活性成分的組成構造明確且能單獨穩定存在。
3. 此活性成分可以工業化規格量產。
4. 此活性成分可以被標準檢測方法分析、定性與定量。
5. 此活性成分有直接科學文獻證明其功能。
6. 此活性成分有直接科學證據顯示其作為食品原料的安全性。
〔臺大教授許瑞祥專文〕你吃的靈芝屬於哪個世代?細說靈芝產業1.0~4.0完整版(上)
〔臺大教授許瑞祥專文〕你吃的靈芝屬於哪個世代?細說靈芝產業1.0~4.0完整版(下)
 Publication Date: Oct 3 2023Author: Ruey-Shyang Hseu, Michael Lee Publisher: Wu Ting Yao Studio, Taipei,...
Publication Date: Oct 3 2023Author: Ruey-Shyang Hseu, Michael Lee Publisher: Wu Ting Yao Studio, Taipei,...
COPYRIGHTS © Ganodermanews SINCE By 2015 ALL RIGHT RESERVED.
靈芝新聞網 版權所有 | 本網站內容作者享有其著作權,禁止侵害,違者必究
Powered by
ganodermanews.com
 最新期/2023年秋季號
最新期/2023年秋季號